La découverte de l’épissage génétique
Le prix Nobel de Physiologie ou Médecine de 1993 fut attribué à Richard Roberts et Phillip Sharp pour leur travaux sur le réarrangement des gènes et l’épissage génétique, un des principes fondamentaux façonnant la diversité protéique et l’évolution.
En 1972, Richard Roberts commença un travail fondamental sur la régulation génétique et la structure de l’Adénovirus qui déboula sur la découverte en 1977 de l’épissage génétique. Assez remarquablement, Richard Roberts et Phillip Sharp ont tous deux indépendamment au même moment et avec le même model et techniques, démontré la théorie du gène fragmenté : une partie du matériel génétique est retiré de l’ARN avant qu’il ne soit traduit en protéines.
Dans les cellules eucaryotes, l’ADN est transcrit en ARNm dans le noyau. Du noyau, l’ARNm est sélectivement transporté jusque dans le cytoplasme où il est traduit en protéines. Cette compartimentation du processus de transcription permet un traitement plus approfondi des gènes que dans les procaryotes.
Quand Roberts et Sharp comparèrent l’ARNm dans le noyau et dans le cytoplasme, ils le trouvèrent diffèrent. En mettant en parallèle des segments d’ADN avec les segments d’ARN cytoplasmique correspondant – en formant des hybrides ARN-ADN – ils trouvèrent qu’ils ne s’assemblaient pas complètement. Certaines régions correspondaient alors que d’autre semblaient avoir complétement disparues dans l’ARN cytoplasmique.
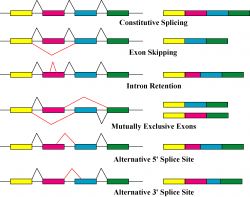
De précèdent travaux ont suggéré que pendant le traitement de l’ARNm, plusieurs régions nucléotidiques sont retirées : les introns. D’autres régions sont assemblées pour former un brin codant pour les protéines : les exons.
Des travaux sur des mutations ont identifiés des ‘séquences consensus’ dans l’ARNm aux sites de reconnaissance, de découpe et d’épissage de la molécule, délimitant les introns des exons et nécessaire pour que l’épissage génétique ait lieu. Ces séquences sont universellement trouvées dans les mammifères, les levures et les plantes. Un tel niveau de conservation permet de dater le processus à des milliards d’années - au moment de l’origine des cellules eucaryotes.
L’épissage génétique peut avoir lieu avec ou sans l’aide d’un splicéosome (une grande machine moléculaire complexe) et différents types d’épissage – cis et trans – ont été identifiés chez les trypanosomes, chez C.elegans et les vertébrés.
Les gènes découpés et recombinés sont très fréquemment trouvés dans les génomes pour deux bonnes raisons. Premièrement, ils permettent facilement à de nouvelles combinaisons génétiques d’apparaitre spontanément par duplication des exons, créant de nouveaux gènes ou protéines. Par exemple, un exon contenant un site actif d’une enzyme peut être dupliqué, déplacé et associé avec une fonction toute aussi importante sur un autre exon, créant une toute nouvelle protéine – un peu comme des blocs de construction. Deuxièmement, un gène peut donc coder pour différentes protéines. Si un gène contient 8 exons par exemple, il peut tout aussi bien faire une protéine en assemblant les exons 1,4 et 6 ou alors en associant les exons 2,4,6 et 7. Deux molécules complétement différentes résulteront de chacune des combinaisons.
Ceci est une des raisons pour laquelle il n’y a pas de corrélation directe entre la diversité génique et la diversité protéique.
Last edited: 10 November 2014 12:49